Research
Elevational Generalism
Most Andean birds inhabit narrow elevational
ranges thought to be set by a combination of biological and environmental factors. Very few species
inhabit more than 3,000 meters in
elevation. Those that do must compensate for large shifts in
temperature, UV exposure, and the partial pressure of oxygen. Are these exceptional species
physiologically adapting in the face of gene flow, or are plastic and behavioral
responses, allowing them to persist across diverse environmental conditions and preventing them from
specializing on narrower elevation zones?
In Gadek et al. 2017 I found that elevational generalists are either in the process of
diversifying, expanding across
the gradient, or undertaking seasonal or resource-pulse-driven elevational migration, and that
elevational generalism is an unstable and transient condition.
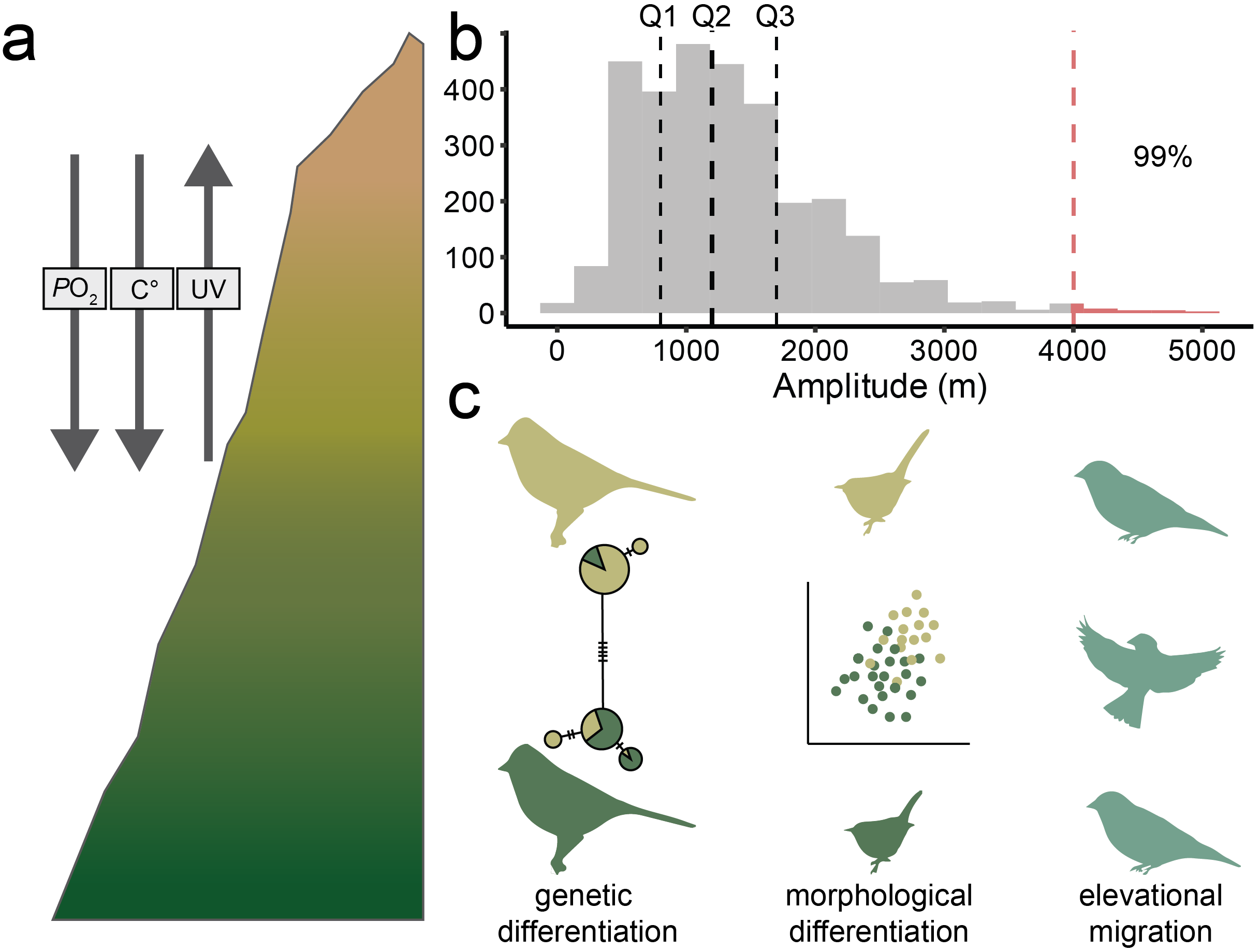
Conceptual figure illustrating the three main mechanisms at play in maintaining or eroding elevational generalist's wide ranges.
Evolution of Elevational Ranges
The evolutionary history of neotropical birds has
undoubtedly been influenced by
Andean uplift, leaving us with current patterns of high diversity and endemism in and around the
Andes. These species' modern elevational ranges represent cumulative histories of expansions and contractions in mountains over evolutionary timescales.
To anticipate how elevational ranges will respond to climate change in the near term, we must understand how they evolved. To this end, I ask: How did elevational ranges
form and change over time? How rapidly have they shifted? Are upward and downward shifts
symmetrically distributed across the
elevation gradient? And how do patterns of range evolution differ among clades?
This work is being done in collaboration
with Eli Stone and Selina
Bauernfeind.
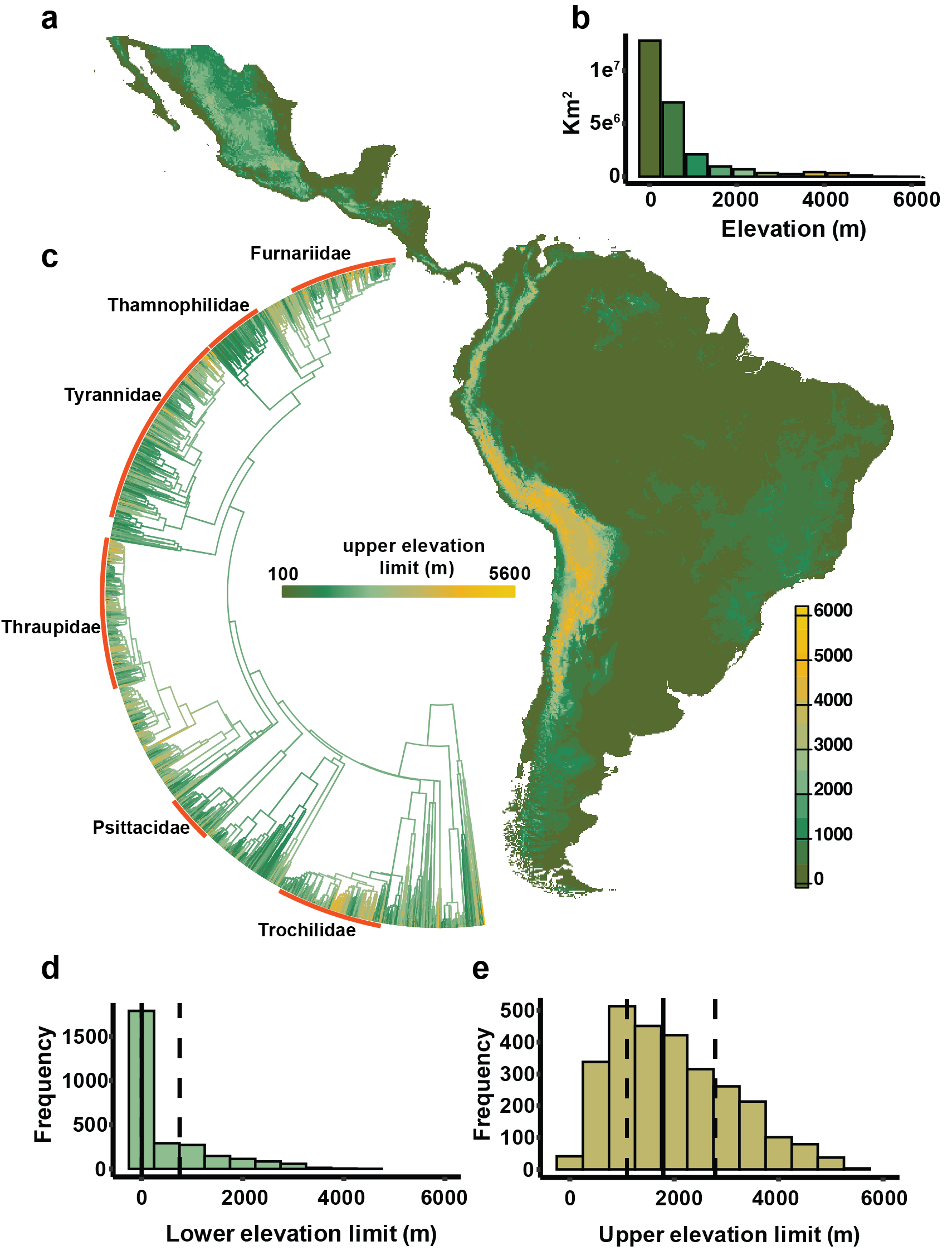
Study context describing elevational ranges of neotropical birds. (a) map of the study region with an elevation model at 30-second resolution. (b) Land area across the study area in 500-m elevation bins. (c) Upper elevation limits of 2,774 species in this study mapped onto a phylogeny (birdtree.org); avian families of over 100 species are highlighted in red. (d) Lower elevation limits and (e) upper elevation limits of the 2,774 species with median (solid bar) and 25% and 75% quantiles (dashed bars).
Plasticity and Adaptation During Extreme Elevational Transitions
Plastic responses to acute changes in elevation are
well documented and shared across many vertebrates.
Similarly, genetic adaptation to high-elevation environments often acts on predictable physiological
pathways. But how do these processes interact, and how do they transition between one another?
To ask these questions, we sampled the entire Peruvian range of two co-distributed marsh birds (
Phleocryptes melanops & Tachuris
rubrigastra). These birds have disjunct elevational ranges,
inhabiting coastal marshes at sea level and high-elevation marshes above 4000 meters. Are the high
elevation populations adapted(ing) to the dramatically lower oxygen availability? Or has gene flow
and recent colonization failed to suppress short-term acclimatization responses? This work is in
collaboration with Jessie
Williamson.

Photograph of Many-colored Rush Tyrant (Tachuris rubrigastra) from Puerto Viejo, Peru, in 2016.
Malaria in Sky Islands
Avian malaria is a widespread chronic disease of birds
caused
by
multiple
Apicomplexan organisms.
To understand how host and pathogen communities vary across the landscape, we surveyed birds and
parasite
communities among
Southwestern sky islands and simulated null communities to compare against empirical data.
We
found that parasite communities differed between sky islands relative to null community models,
suggesting idiosyncratic colonization and extirpation dynamics. This work is in collaboration with Christopher
Witt,
Lisa Barrow, Jessie Williamson, Selina
Bauernfeind, and Rosario
Marroquin-Flores.
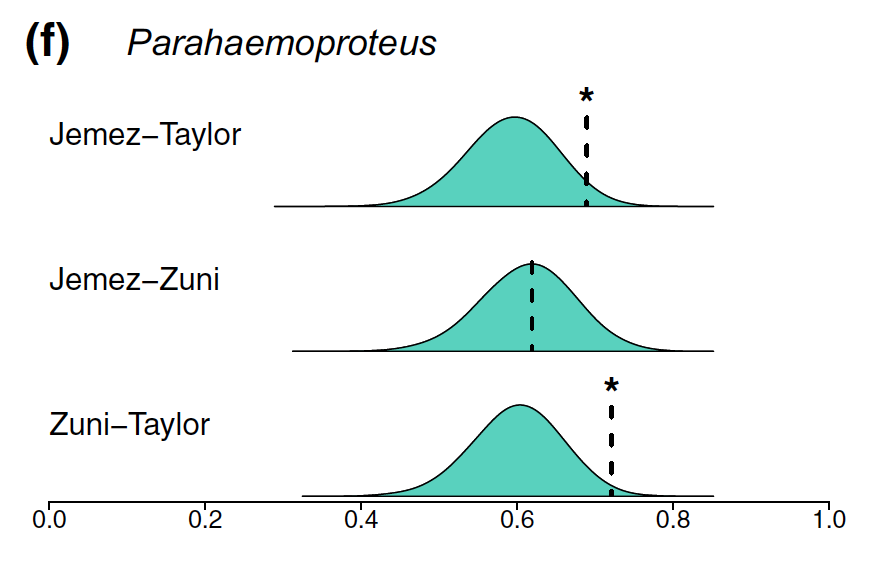
Figure modified from Barrow et al. 2021 showing observed (dashed lines) and expected Jaccard Index values. Expected values obtained from 10,000 randomly simulated communities. Illustrates how observed communities in two "sky islands" differ from expectations.
Tracheal Evolution in Sandhill Cranes
The sandhill crane (Grus canadensis ) is among the few bird species that exhibit tracheal elongation. Within this species, there is substantial size dimorphism between subspecies. Jones & Witt 2014 found that the smaller subspecies that undertake longer distance migrations had proportionally longer trachea hypothesized to make smaller birds sound bigger. I am interested in assessing the symmetry and strength of sexual dimorphism within sandhill cranes.

Photo of sandhill crane trachea coiled within the bird's sternum.
Evolutionary and ecological drivers of microbe-host associations in the lung mycobiomes of birds
Microbiomes are being published at a rapid pace. Yet
wild
bird microbiomes and specifically their fungal components
(mycobiomes) are largely undescribed. Furthermore, how microbiomes are structured by phylogenetic, geographic, and life history traits
remains understudied.
Baseline knowledge of the makeup of these communities and factors that affect their assembly and
maintenance holds important implications for disease ecology, public health, and large-scale
coevolutionary processes.
We are describing the first lung mycobiomes of birds
and modeling the associations of phylogeny, morphology, and ecology with lung fungal communities. This
project is in collaboration with Paris
Hamm & Michael
Mann utilizing samples collected and stored in the Museum of Southwestern Biology.
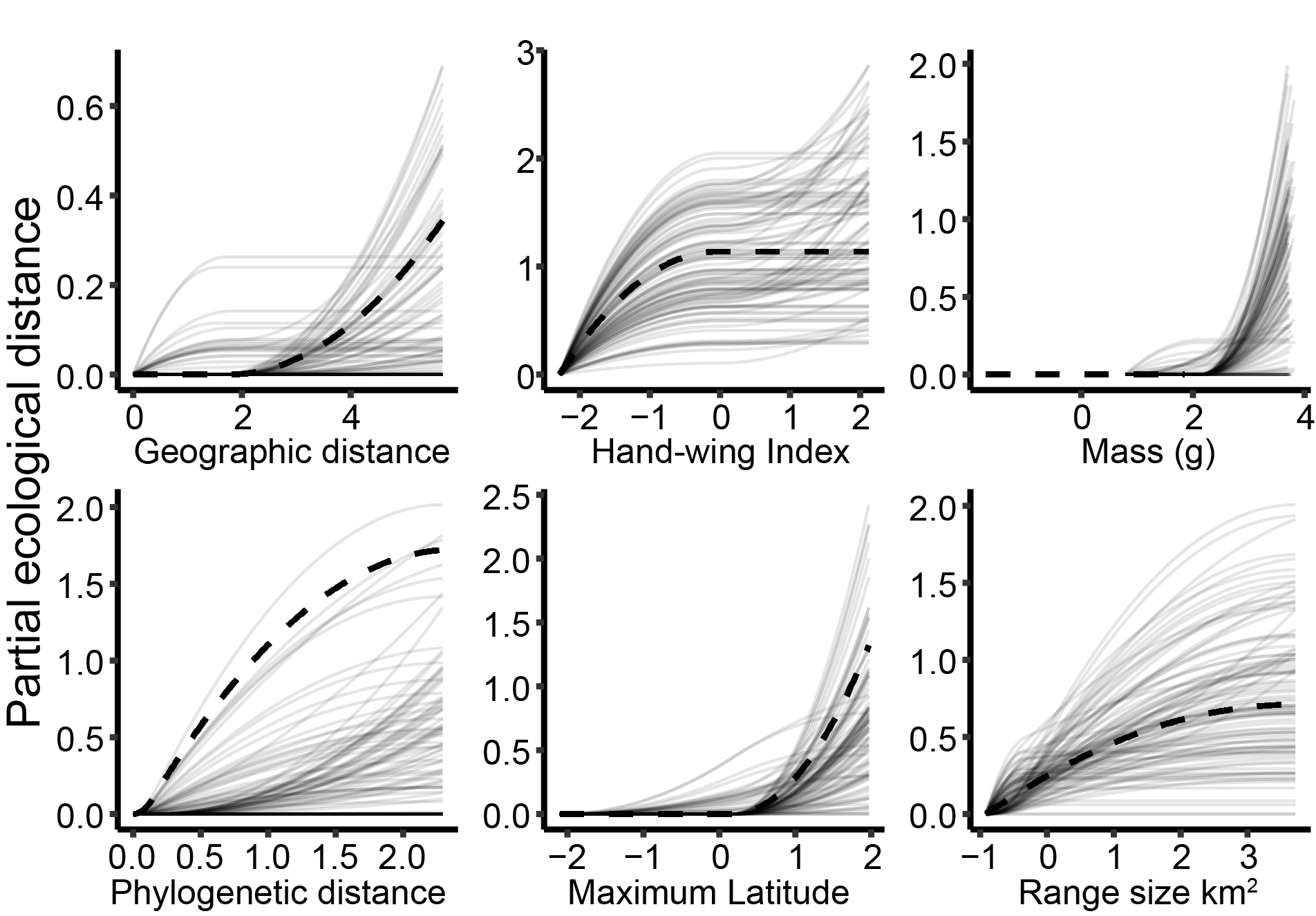
Global (dashed lines) and subsampled (solid lines) generalized dissimilarity models showing geographic, morphological, and phylogenetic predictors of fungal community dissimilarity. Figure shows (height of lines) that phylogenetic distance, hand-wing index, and maximum latitude of bird hosts explain the most dissimilarity among lung fungal communities